Chris Henggeler of Kachana Pastoral Company compiled the paper below whilst attending a Soil Biochemistry Course hosted by Southern Cross University, Lismore, Australia.
We would like to extend our sincere appreciation and thanks to the EJ Connellan Trust for sponsoring a significant portion of Chris� travel costs, thus enabling him to further his education and understanding in Natural Resource Management.
�We desperately need to complement our practical skills and experience with up-to-date scientific knowledge, if we wish to make the responsibilities of Natural Resource Management attractive to the next generation. - Handing over deteriorating resources with an explanation of what went wrong is simply not good enough; we need to offer our heirs direction and a way forward that offers hope and reward.
�Private agriculture has come under much pressure in recent years. We face challenges like genetic contamination, water-shortages, peak oil and shifting market forces� It is exciting to know there are scientifically sound and safe options to address all of these issues.
�The support by the EJ Conellan Trust not only enabled me to directly access cutting-edge scientific information, it also enabled me to mix with other forward thinking people from other parts of the country. I believe the value of this experience to be ongoing.�
The EJ Connellan Trust, provides a range of assistance to people living in remote areas. There is individual assistance for people with special education needs, and there's the actual award itself. The Trust was the brainchild of E J Connellan (EJ) who established Connellan Airways in 1939. Please find out more about the trust on the website: http://www.connellanairwaystrust.org.au/about.php.
For more detailed information about Soil Fooweb Inc. please visit their website: www.Soilfoodweb.com
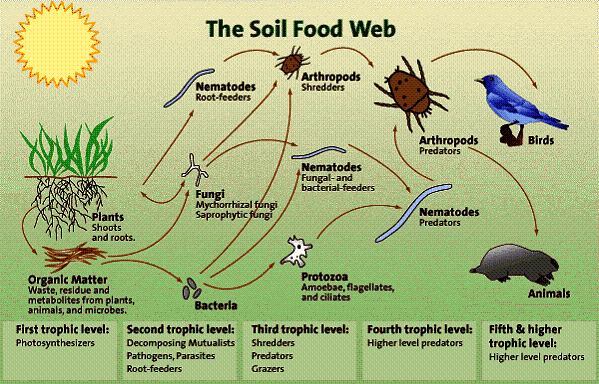
Soil food-web structure and function in a sub-catchment typical for northern Australian rangelands.
July 2006: An interpretation by Chris Henggeler based on over twenty years of observation in the field and the comparing of data provided by Soil Foodweb testing since 2002. Figure 1.
Figure 1.
Landscape typical for many upper-river rainfall-catchment areas in northern Australian rangelands.
In this case:
Rangeland implies the absence of intensive management/land-use. In general such areas are considered to have low or limited productive value. They have no history of chemical contamination or mechanical disturbance. Much of the rangeland embraces areas that are located beyond the immediate spheres of modern human land-use.
Sub-catchment implies that the available source of water is what drops out of the sky and/or water from earlier rainfall that has been retained on or below the surface.
In past times much of Northern Australia was covered in rain-forest (Flannery et al.). Areas that today are shallow basins in upper-river rainfall-catchment areas would then have served as soaks and sponges, gradually releasing water into down-stream drainage lines and feeding perennially flowing river systems draining either inland or towards the ocean.
Climax communities would have been supported by a resilient and complex soil foodweb very similar to that depicted in ‘figure 2’ below. Similarly complex food-webs would have existed in the litter (compost foodweb) and at limb and foliar levels (foliar foodweb). (Visit the SFI website for images and diagrams: www.soilfoodweb.com )
A vast and diverse range of animal species at higher trophic levels would have assisted with functions like:
- aeration of soils (burrowing animals)
- perennial cycling of vegetation (eating and digesting plant material)
- trampling and crushing litter (making the job easer for shredders, fungi and bacteria)
- tearing down live and dead limbs (adding to the nutrient supply of recyclers)
- creating disturbances in micro-environments
- creating habitat and edge effect at the microenvironment levels
- serving as “taxi-cabs” and agents of biological inoculations (seeds, micro-organisms).
(Flannery suggests that in recent millennia Australia may have lost as much as 95% of its mega-faunal species; there are no estimates of the according amount of active animal biomass; little is known about the ripple effect that these extinctions had on species and functional groups of species at lower trophic levels.)
 Figure 2.
Figure 2.
Vigorous tropical plant growth throughout upper, middle and lower storey vegetation would have allowed for efficient capture of the daily supply of solar energy via photosynthesises to fuel a system that would have been trapping and storing “sunlight- energy” largely in the form of active biomass; a gradual net-accumulation of organic carbon would have been possible. Mosses and algae would also have thrived in the resulting microclimate. (Trials are currently being conducted to mimic such processes on Kachana Station with SFW data being collected every five years:
http://www.kachana.com/workshops_education/05_klmw_soil_profile_8.php
http://www.kachana.com/workshops_education/05_klmw_soil_profile_6.php)
100% ground-cover would have protected soil-surfaces from the pounding force of tropical rainfall and enhanced the recycling of water many times over each season. Raindrops would have been shattered and the water slowed down before being rapidly absorbed into the soil profile. Excess water would have gradually percolated deeper into the ground feeding aquifers and sponges; some water may have been released laterally into drainage lines. Good soil structure would have ensured sufficient moisture to sustain high levels of soil-microbial activity even during low-rainfall periods. Transpiration by vegetation would have maintained a humid microclimate (perhaps even being conducive in prolonging convectional rainfall activity). There would have been sufficient soil-surface humidity for vigorous microbial activity to be sustained throughout the year.
Carbon and mineral cycling would have been continuous throughout the year as a result of species from several trophic levels constantly interacting.
The impact of both internal and external disturbances would have been significantly buffered by the forest microclimate below an upper- or middle-story canopy. The complexity of the food-web at all levels and the amount of energy that would have been stored in the form of biomass would indicate an extremely productive and resilient eco-system.
In general disturbances would have been limited to storms and low frequency fires by lightning. Ignition would have occurred late in the dry season or early during the wet season. Seasonal variation and animal response (to the season) would have determined the intensity, and how extensively a fire would have burned.
Limiting factors would have included:
- microclimate (high humidity and dew)
- effectiveness of the carbon cycle (according absence of litter already incorporated in soil or new life)
- low surface temperatures
- rising moisture levels in soil and litter (try lighting a camp fire on a cold damp morning)
- growth phase of the plant (healthy growing plants will not burn readily and tend to slow down a fire)
- type of vegetation (broad-leafed forest vegetation generally does not burn readily)
- rainfall (halting spread)
- healthy riparian zones (limiting spread)
Intensity patterns within burned areas would have contributed to habitat diversity and therefore biodiversity: ignition areas having experienced the most intense heat, the intensity then petering out towards the edges and allowing for a “cooler” burn in many places (personal observation of lightning-fires and burning in managed country: http://www.kachana.com/projects_research/or_fire_experiment.php).
Wide-spread high intensity fires would have been a rare event leaving much time for recovery and recolonisation.
Coinciding with the advent of humans to Australia much has changed in recent millennia. The way in which species loss occurred and the impact of new species remains the subject of debate. It is however safe to say that a complex array of compounding factors did exist, some of which have been studied. (Kohen, Flannery, Savory, Diamond et al.)
Today we deal with lower/low levels of succession in above and below ground communities. Arguably we are now faced with new “exotic” environments harbouring a variable mix of remnant native species and recent arrivals to the continent. These are now jointly exposed to an unprecedented combination of disturbances.
We observe new fire sub-climaxes determined by the intensity and frequency of human-lit fire every three to five years. In some areas soil-loss is matched by crumbling and breaking down of surface materials giving an impression of landscape stability.
Our typical rangeland sub-catchment has changed in appearance and it could be argued that eco-system processes are now largely dysfunctional and each area would have its own story to tell.
(Eco-system function: http://managingwholes.com/--ecosystem.htm )
Only a fraction of former levels of sunlight energy are now entering the eco-system via photosynthesis to later become available to higher trophic levels. Much of the sun’s energy is merely heating up the ground and then radiated back contributing to the drying out of the soil-surface and modifying of microclimates.
Rainfall is seasonal and erratic and averages across Northern Australia range between 200 and 900 mm per annum. Even in high rainfall areas, not much water is retained at the locations where precipitation occurs. This leads to poor and ineffective water-cycles (http://managingwholes.com/eco-water-cycle.htm). Compaction, wind-, raindrop- and algal-capping severely restrict the absorption of water into the ground. Water that does not run off as a flood is then largely lost to surface evaporation. Only a fraction becomes available to support biological activity on site. High rainfall events are often associated with erosion and down-stream flooding: http://www.kachana.com/photo_gallery/photos_disturbances_water.php
(Further study needs to go into the limiting nature of ineffective rainfall and its cost to biological and commercial productivity. Storey: http://www.kachana.com/projects_research/or_a_storey_waterways.php )
The cycling of minerals mostly takes place within the soil. Above the ground small reptiles and small marsupials may reside fully within such a sub-catchment; non-resident larger animals and birds would at times contribute to the cycling or transfer of nutrients. Above ground dead organic matter is subjected to solar radiation and oxidation for much of the year. Water-soluble minerals are brought to the surface as moisture evaporates off exposed soils. Where there are no crusts to hold things in place we observe net transfer of minerals out of the landscape as a result of fire, wind- and water-erosion. (Fire will of course also reduce the biomass and alter the nature of algal capping.)
Biological communities remain highly complex, but their resilience has been greatly altered. Little vegetation survives to serve the role of supplying fuel to the food-web and the genetic viability of animals at higher trophic levels is questionable. Both structure and function of the remaining (modified and adapted) soil food-web is now limited by the level of sunlight capture of the remaining vegetation in these areas. With 80% to 90% of the soil surface being exposed to solar radiation, wild temperature fluctuations and seasonal torrential downpours, vegetation is limited to scattered mature trees (upper storey), some shrubs and predominantly annual grasses and “weeds”. The sandy surfaces in-between are held together by wind- and raindrop-capping and a thin algal crust. In general we need to consider succession having been suppressed to levels that reflect 'fungal : bacterial' ratios in the vicinity of 1:10.
(See: http://www.soilfoodweb.com.au/index.php?pageid=328; At present there is not enough North-Australian data available to support this supposition.)
Given that most of the “soil” in bare areas (beneath the algal crust) is sand, we must suspect that there is no soil-foodweb as such that supports the whole sub-catchment. Rather we may be seeing a range of soil-food-webs limited to various sites/locations:
Algal crust; annuals and weeds; rhizospheres of individual isolated bushes; rhizospheres of individual isolated trees; location specific micro-environments like the insides of dead/dying trees; under loose stones and rubble (where there is some physical protection from direct exposure to the prevailing forms of disturbance).
Funding is being sought to conduct soil foodweb sampling of these micro-environments. Available data shows the existence of members of the required functional groups, but we do not know enough about their concentrations and distribution within the soil. It is possible that species composition may vary considerably between sites; this will depend on root patterns of shrubs and trees and whether or not we have mycorrhizal colonisation. Meanwhile the best we can do is guess what is happening based on available data from other areas (http://www.kachana.com/projects_research/mp_soilfoodweb.php ):
Algal Crusts: Biological activity is largely seasonal; most organisms would be dormant for much of the year.
F:B <1:10 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Algae - bacteria & fungi – protozoa – bacteria – algae
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
Limited soil-building 1mm to 2mm beneath crust may allow for nematode activity and the occasional presence of higher level predators. These crusts will also grow on rock surfaces, gradually breaking them down. Systems seem to be quite stable despite their lack of productivity when viewed from a commercial perspective.
Bare ground, annuals and weeds: Biological activity is largely seasonal; most organisms would be dormant for much of the year. We readily observe the removal of surface organic matter by ants, wind, fire or flood.
F:B = 1:10 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Plants - bacteria & fungi – protozoa, nematodes, micro-arthropods –predatory nematodes (likely) and larger arthropods – small reptiles
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
Rhizospheres of individual isolated bushes: Biological activity is maintained throughout the year, but slows down significantly during the drier months.
F:B = 3:1 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Plants - bacteria & fungi – protozoa, nematodes, micro-arthropods –predatory nematodes (likely) and larger arthropods – small reptiles – larger herbivores.
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
We must suspect a diverse range of species and the presence of all functional groups. We suspect that productivity and advancing succession is limited by fire.
Rhizospheres of individual isolated trees: Biological activity is maintained throughout the year, but slows down significantly during the drier months.
F:B = 300:1 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Plants - bacteria & fungi – protozoa, nematodes, micro-arthropods –predatory nematodes (likely) and larger arthropods – small reptiles
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
Here we suspect a self-regulating pattern that limits succession: Frequent burning of litter and mulch prevents the “composting” organisms from developing into fully functional groups so the nutrient uptake for the tree may be largely dependent on the biological processes involved in the break-down of parent material (chelation) and the “mining” of minerals. Flannery quotes literature indicating that trees will produce toxins that will reduce competition for nutrients.
Insides of dead/dying trees: Biological activity is maintained throughout the year, but slows down significantly during the drier months.
F:B = 500:1 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Plants - bacteria & fungi – protozoa, nematodes, micro-arthropods –predatory nematodes (likely) and larger arthropods – small reptiles.
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
Soil has a good smell to it and is well aggregated; hyphae are visible. This could be an indicator that the range of organisms required for a functional foodweb across the whole sub-catchment may be present, but these lack the conditions to expand.
Under loose stones and rubble: Most biological activity takes place below the ground and is maintained throughout the year, but slows down significantly during the drier months. Roots are physically protected from fire, heat and compaction by rocks. Below these protected areas we find aggregated sand, silt and clay and lower sides of rocks show signs of weathering. This too could be an indicator that the range of organisms required for a functional foodweb may be present. These are the foodwebs that need to be tested for presence of important functional groups before we consider the introduction of species from other areas. Given that there is no artificial chemical history in these soils it is likely that mycorrhizal fungi will be present.
F:B = 3:4 ? (http://www.soilfoodweb.com.au/index.php?pageid=328)
Plants - bacteria & fungi – protozoa, nematodes, micro-arthropods –predatory nematodes (likely) and larger arthropods – small reptiles
(Relationships as in the diagram on page 5 of the Soil Biology Primer or http://www.soilfoodweb.com.au/index.php?pageid=323 )
 Figure 3.
Figure 3.
The removal of a rock exposes soil and root structure at the base of a perennial grass plant in an area that had recently been subjected to fire. Human lit fires are arguably a biological disturbance; as are human induced floods that result from the way we manage our upper-river rainfall-catchments. http://www.kachana.com/photo_gallery/photos_disturbances_fire.php
http://www.kachana.com/photo_gallery/photos_disturbances_water.php
http://www.kachana.com/environmental_management/pp_prepare_for_rain.php
In ‘Figure 3.’ above we notice how fire would have removed dead leaf material from a senescent perennial grass prompting fresh growth by bringing light to the growth points and by re-activating below ground soil foodweb function.

Conclusion:
In much of northern Australia past and current human induced disturbances are now the factors that limit biological succession above and below our soil-surfaces. In many instances we may expect the soil-foodweb to be structurally intact, yet severely weakened or at risk due to inappropriate soil-surface management. It may be possible to achieve significant increases of productivity simply by introducing practices that will enhance existing soil foodweb function and favour the particular functional groups that are relevant to the production expectations of an area.
Chris Henggeler, Kachana Pastoral Company, www.kachana.com
July 2006
Sample of reclamation work in progress on Kachana Station:
What we started off with in 1992…

The same area a few years later:
 |
|
|
|


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Soil profile information of this area:
http://www.kachana.com/workshops_education/05_klmw_soil_profile_8.php
Other related links and web-sites:
Beyond Conventional Pastoralism
Flash animation of the water cycle (850 K).
Allan Savory Africa Report June 06
Erosion: why it happens and what to do about it
There is no quicker way to sequester carbon than to manage for healthy savannahs
http://www.soildynamics.com/living.htm
http://www.soildynamics.com/erosion.htm
“Hot air in Northern Australia – taking the red-steer by the horns.”
top
|
Last modified: October 22 2019 23:27:42. |
![]() print this page
print this page